

1. Подготовка к вопросам по теме Цитология
.docxТема: «ЦИТОЛОГИЯ (биология клетки)»
Основные вопросы темы:
Общий принцип организации эукариотической клетки.


Общий принцип строения клеточной оболочки – плазмолеммы.
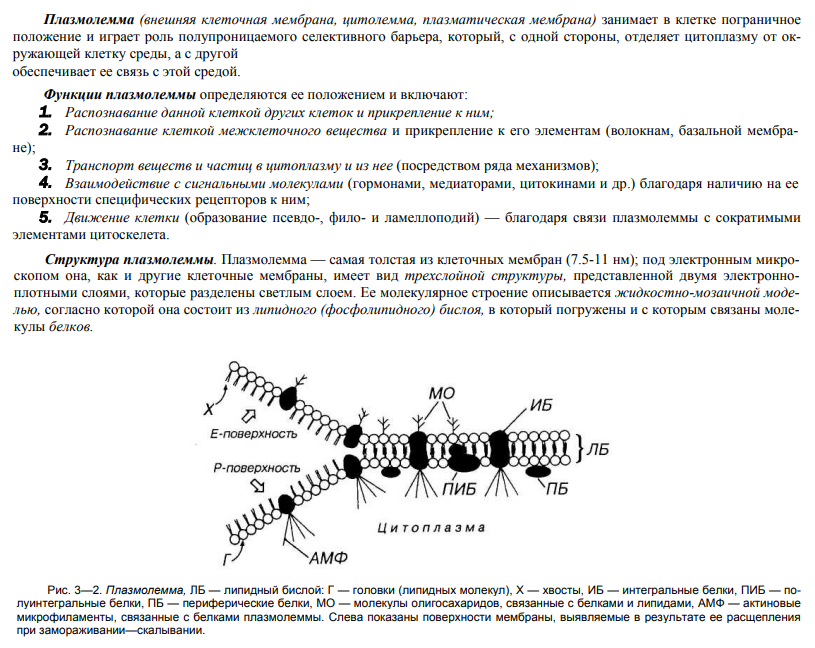


Строение биологической мембраны. Жидкостно-мозаичная модель мембраны.
В 1972 г. Сингер и Николсон (Singer, Nicolson) предложили жидкостно-мозаичную модель мембраны, согласно которой белковые молекулы плавают в жидком фосфолипидном бислое. Они образуют в нем как бы своеобразную мозаику, но поскольку бислой этот жидкий, то и сам мозаичный узор не жестко фиксирован; белки могут менять в нем свое положение. Покрывающая клетку тонкая мембрана напоминает пленку мыльного пузыря — она тоже все время «переливается».
На рисунке представлено плоскостное изображение жидкостно-мозаичной модели мембраны и ее трехмерная модель.
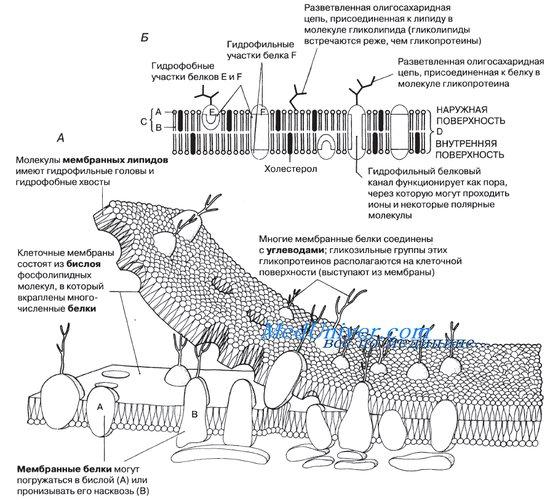
Ниже суммированы известные нам данные, касающиеся строения и свойств клеточных мембран.
1. Толщина мембрансоставляет около 7 нм.
2. Основная структура мембраны— фосфолипидный бислой.
3. Гидрофильные головы фосфолипидных молекулобращены наружу — в сторону водного содержимого клетки и в сторону наружной водной среды.
4. Гидрофобные хвостыобращены внутрь — они образуют гидрофобную внутреннюю часть бислоя.
5. Фосфолипидынаходятся в жидком состоянии и быстро диффундируют внутрибислоя— перемещаются в латеральном направлении.
6. Жирные кислоты, образующие хвосты фосфолипидных молекул, бывают насыщенными и ненасыщенными. В ненасыщенных кислотах имеются изломы, что делает упаковку бислоя более рыхлой. Следовательно, чем больше степень ненасыщенности, тем более жидкую консистенцию имеет мембрана.
7. Большая часть белков плавает в жидком фосфолипидном бислое, образуя в нем своеобразную мозаику, постоянно меняющую свой узор.
8. Белки сохраняют связь с мембраной, поскольку в них есть участки, состоящие из гидрофобных аминокислот, взаимодействующих с гидрофобными хвостами фосфо-липидов; вода из этих мест выталкивается. Другие участки белков гидрофильны. Они обращены либо к окружению клетки, либо к ее содержимому, т. е. к водной среде.
9. Некоторые мембранные белкилишь частично погружены в фосфолипидный бислой, тогда как другие пронизывают его насквозь.
10. К некоторым белкам и липидам присоединены разветвленные олигосахаридные цепочки, играющие роль антенн. Такие соединения называются соответственно гликопротеинами и гликолипидами.
11. В мембранах содержится также холестерол. Подобно ненасыщенным жирным кислотам он нарушает плотную упаковку фосфолипидов и делает их более жидкими. Это важно для организмов, живущих в холодной среде, где мембраны могли бы затвердевать. Холестерол делает мембраны также более гибкими и вместе с тем более прочными. Без него они бы легко разрывались.
12. Две стороны мембраны, наружная и внутренняя, различаются и по составу, и по функциям.
5) Химический состав мембран. С помощью световой и электронной микроскопии в клетках выявлены разнообразные мембранные структуры. Все они имеют сходный химический состав и принцип организации, но в зависимости от типа мембран и их функций соотношение химических компонентов и детали строения могут отличаться.
Мембраны состоят из липидов, белков и углеводов (рис.16). Липиды составляют в среднем 40% сухой массы мембран. Среди них преобладают фосфолипиды (до 80%).
Основным функциональным компонентом биологических мембран являются белки. Но только образовав прочные комплексы с липидами, они способны проявлять активность.
Поверхностные белки (около 30% от общего количества мембранных белков) размещены на наружной и внутренней поверхностях мембран и связанные с последними электрическими силами непосредственно или через двухвалентные катионы, преимущественно Са2 + и Mg2 +. Они легко отделяются от мембран после разрушения клеток.
Внутренние белки (почти 70% общего количества мембранных белков) погружены в двойной слой липидов на разную глубину, а в некоторых случаях пересекают мембрану насквозь. Такие белки связывают обе поверхности мембраны.
Углеводы входят в состав мембран не самостоятельно, а образуют комплексы с белками или липидами.
Организация биологических мембран. Сейчас общепринятой является модель растворимо-мозаичной строения мембран (рис.16). Такое название произошло от того факта, что около 30% липидов тесно связаны с внутренними белками, а остальное - находится в жидком состоянии, где «плавают» липопротеиды. Молекулы липидов размещены в виде двойного слоя, их полярные гидрофильные «головки» обращены к внешней и внутренней сторон мембран, а гидрофобные неполярные «хвосты» - внутрь.
Если посмотреть на мембрану сверху, то она напоминает мозаику, созданную полярными «головками» липидов, поверхностными и внутренними белками. Толщина мембран варьирует в довольно широких пределах в зависимости от их типа. Мембраны клеток эукариот и прокариот сходны по строению.
Между молекулами белков или их частями часто существуют поры (канальцы), заполненные водой. Молекулы, входящие в состав мембран, способные перемещаться, благодаря чему мембраны быстро возобновляются за незначительных повреждений, образуются над оголенными участками цитоплазмы, могут легко сливаться друг с другом, растягиваться и сжиматься, например, при движении клеток или изменения их формы.
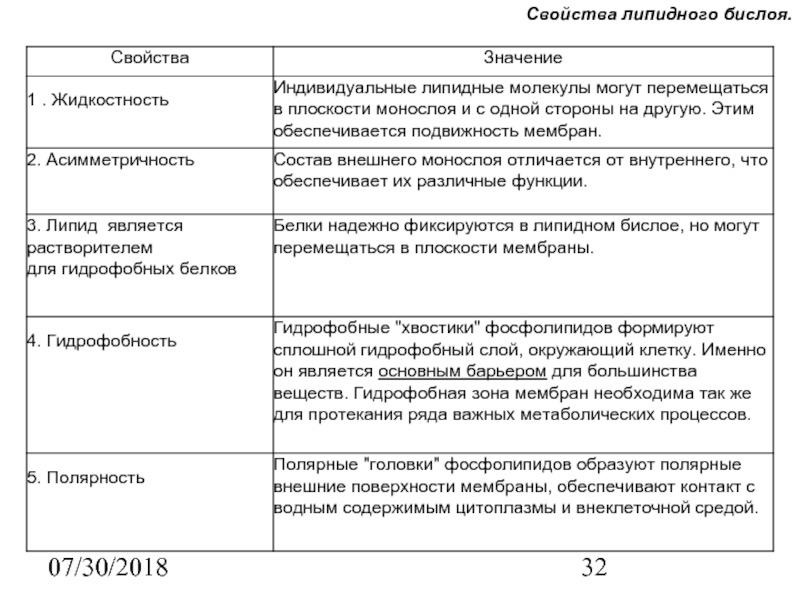
Свойства мембраны, обусловленные наличием «липидного бислоя».
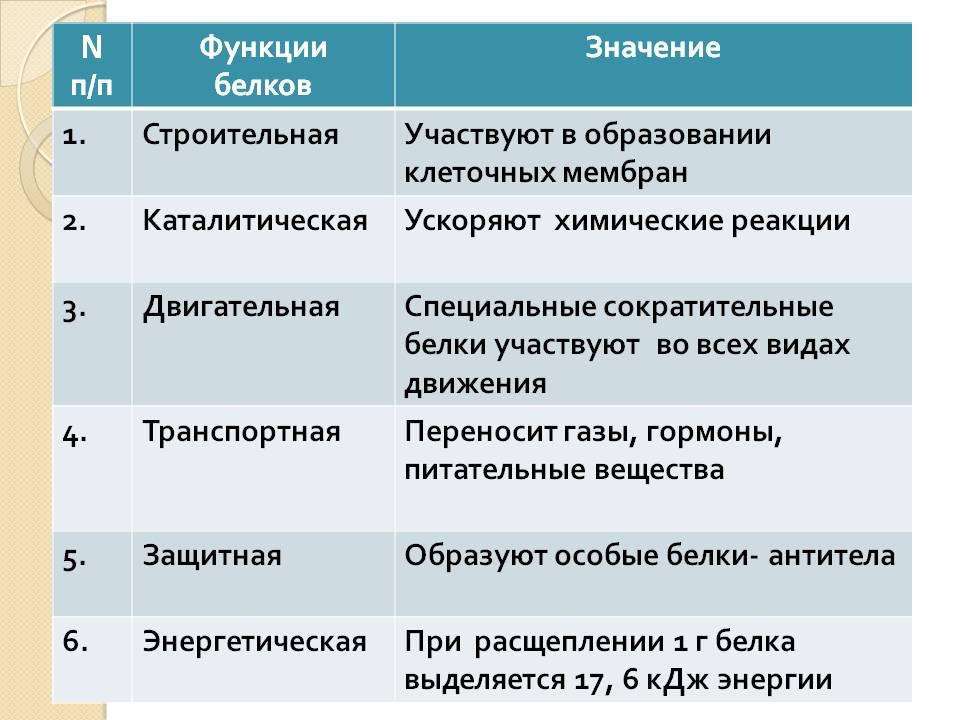
Свойства мембраны, обусловленные наличием белков; биологическая роль белков.

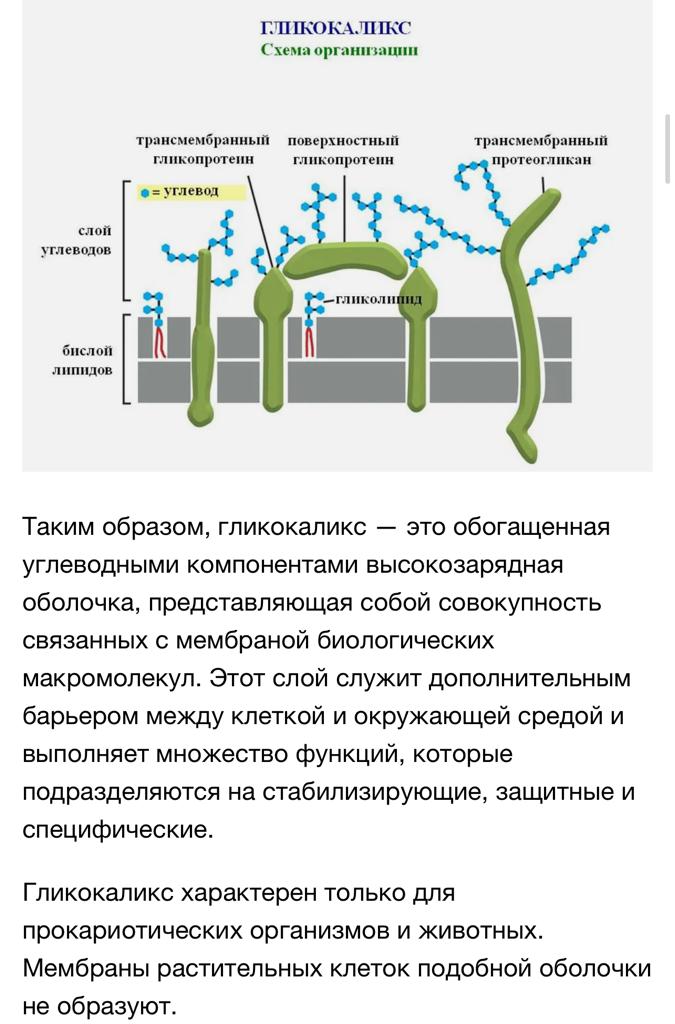
Строение и роль гликокаликса.

Гликокаликс выполняет рецепторную и маркерную функции, а также участвует в обеспечении избирательности транспорта веществ и пристеночном (примембранном) пищеварении.

Строение и роль субмембранного комплекса – кортикального слоя.

Мембранный транспорт: пассивный, активный, облегченный.

Эндоцитоз, его разновидности.
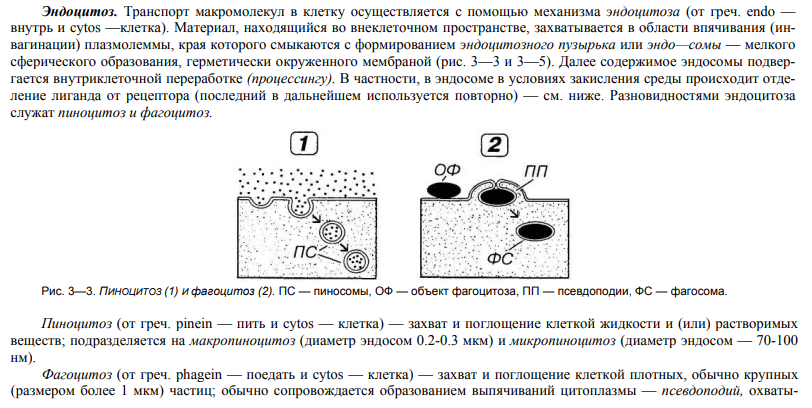

Строение и роль окаймленных ямок и пузырьков.


Понятие об экзоцитозе и трансцитозе.


Понятие о мембранных рецепторах и выполняемые ими функции.

Межклеточные соединения – механические: интердигитации, десмосомы, промежуточные соединения, плотные соединения.
![]()
![]()



Межклеточные - коммуникационные соединения: щелевые, синапсы.
![]()

![]()

Цитоплазма, компоненты её составляющие.
![]()
Понятие о гиалоплазме.

Строение и значение гранулярной эндоплазматической сети.


Строение и значение агранулярной эндоплазматической сети.

Строение и значение комплекса Гольджи.



Строение и значение митохондрий.



Строение и значение лизосом.


Типы лизосом.


Строение и значение пероксисом.


Строение и значение рибосом.


25. Цитоскелет клетки, его структурные элементы и их производные.

26. Строение и значение клеточного центра.
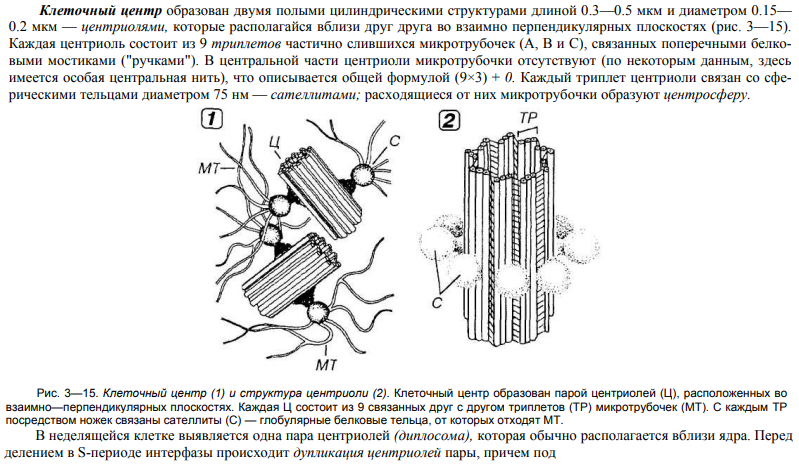

27. Строение и значение микротрубочек.
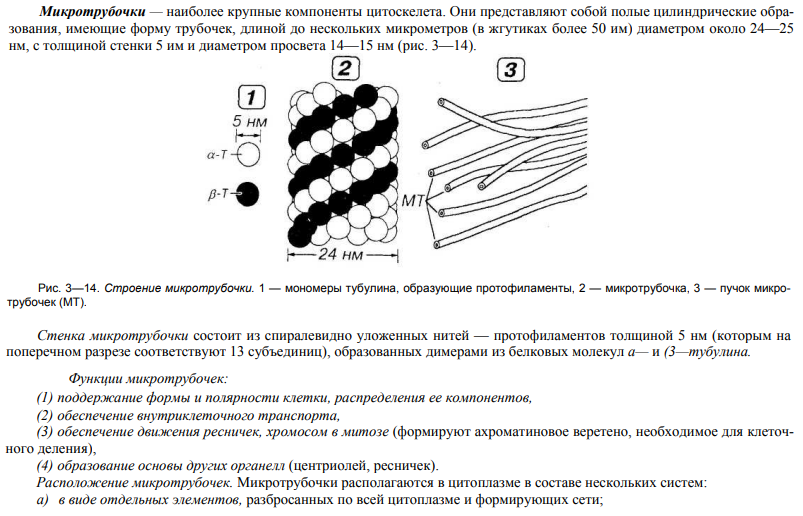

28. Строение и значение микрофиламентов.


29. Строение и значение промежуточных филаментов.


30. Понятие об органеллах специального значения.
Органеллы специального назначения – это постоянно присутствующие и обязательные для отдельных клеток микроструктуры, выполняющие особые функции, которые обеспечивают специализацию ткани и органа. К ним относят: реснички, жгутики, микроворсинки, миофибриллы.
31. Строение и значение мерцательных ресничек.

32. Строение и значение микроворсинок.
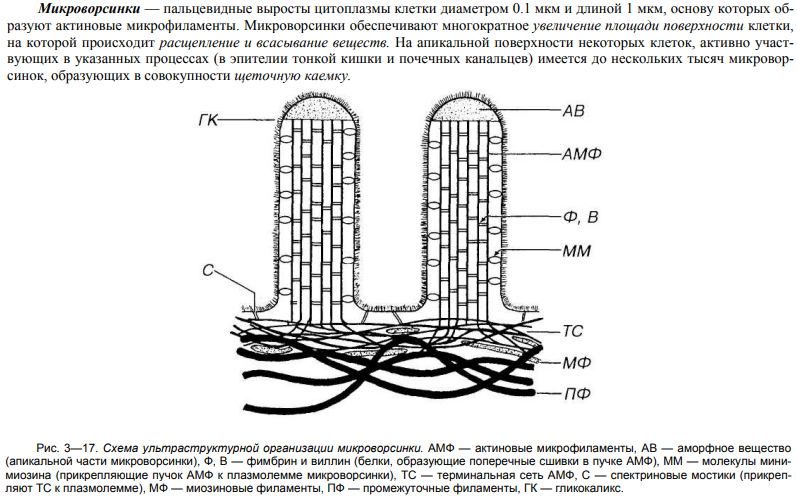
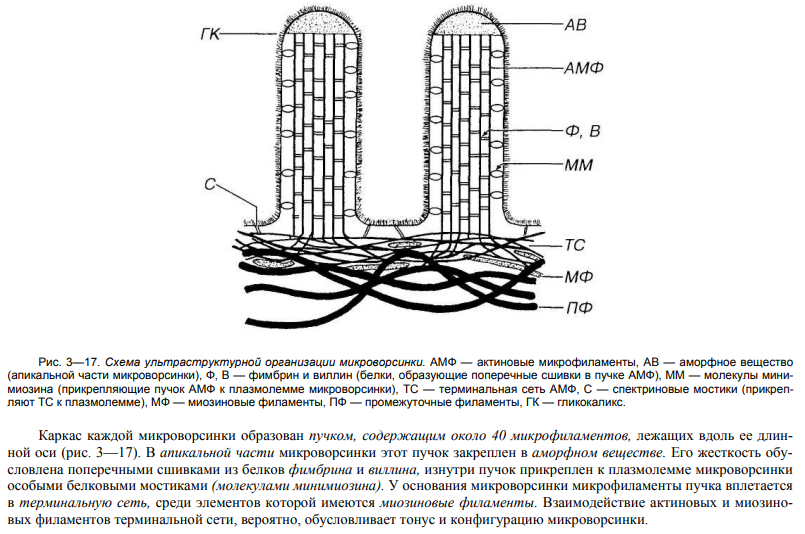
![]()
33. Включения цитоплазмы, их классификация, значение.

34. Ядро клетки, общий план строения и значение.
![]()

35. Строение и значение ядерной оболочки.
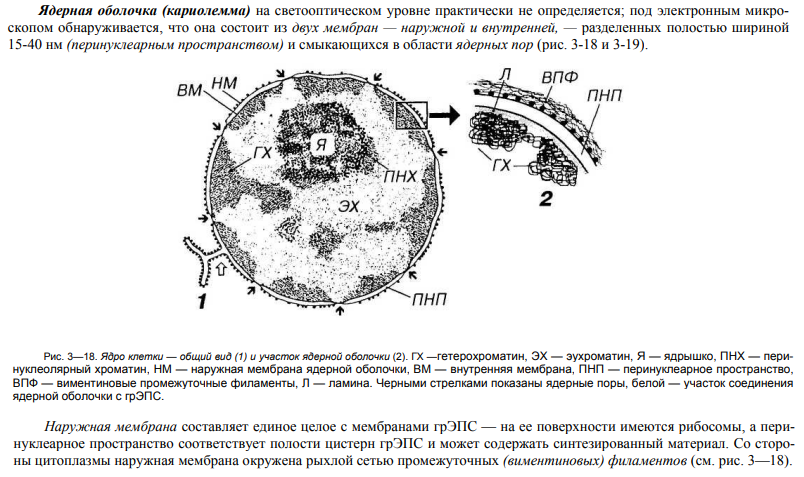

36. Строение и значение хроматина, виды хроматина.




З7. Морфологическая характеристика хромосом.




38. Строение и значение ядрышка.

З9. Понятие о кариоплазме, её состав.

40. Понятие о клеточном цикле, его периоды.


41. Постмитотический (G1) период, происходящие в нем события.

42. Синтетический (S) период. Его значение.

43. Премитотический (G2) период, происходящие в нём события.

44. Митотический (М) период, его общая характеристика, стадии.

45. Характеристика стадий митоза: профазы, метафазы, анафазы, телофазы.
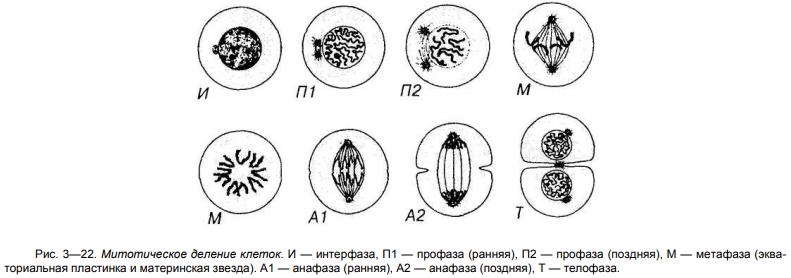

46. Понятие об эндомитозе, полиплоидии, политении.



47. Биологические особенности и значение мейотического деления.
1. Это механизм, поддерживающий постоянство числа хромосом в ряду поколений.
2. При мейозе образуется большое количество различных новых комбинаций негомологичных хромосом, что обеспечивает комбинативную изменчивость.
3. В процессе кроссинговера также происходит рекомбинация генетического материала. В этом одна из причин изменчивости организмов, дающая материал для отбора.
Между митозом и мейозом можно выделить следующие основные различия:
1. Мейоз состоит из 2 последовательных делений, которым предшествует однократная редупликация ДНК. Митоз состоит из одного деления с однократным удвоением ДНК.
2. В профазу I мейоза происходит конъюгация и кроссинговер, чего нет в профазе митоза.
3. Во время мейоза в метафазу I по экватору выстраиваются биваленты, и в анафазу I расходятся гомологичные хромосомы, а во время митоза – хромосомы и сестринские хроматиды соответственно.
4. В телофазу I мейоза не происходит деспирализации хромосом, не образуется ядерной оболочки и ядрышка.
5. В результате мейоза образуются четыре дочерних клетки с гаплоидным набором хромосом, а в результате митоза – две дочерние клетки, содержащие диплоидный набор хромосом.
Процесс формирования половых клеток называется гаметогенезом. Он содержит в себе 4 периода: размножения, роста, созревания, формирования.
48. Понятие о стабильных, растущих и обновляющихся популяциях клеток.

49. Регуляция клеточного цикла: значение протоонкогенов и антионкогенов, факторов роста, кейлонов.
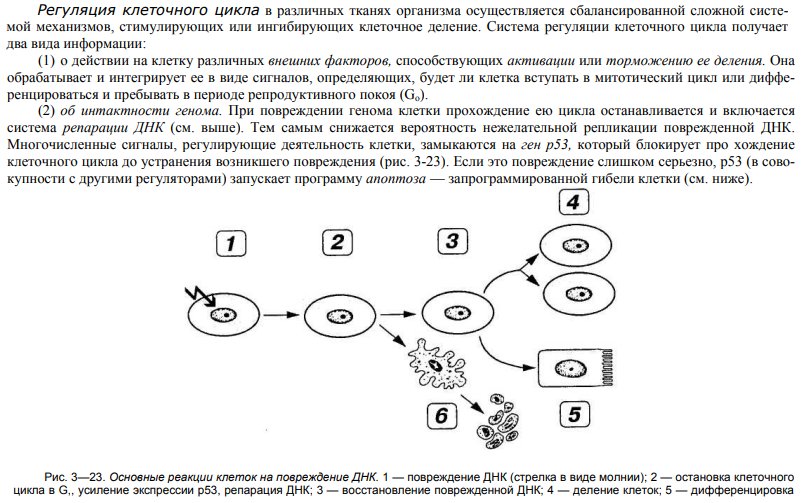


50. Некроз (гибель) клеток – характеристика морфологических изменений.


51. Апоптоз – генетически контролируемая (может быть запрограммированной) гибель клеток. Общие представления, значение.




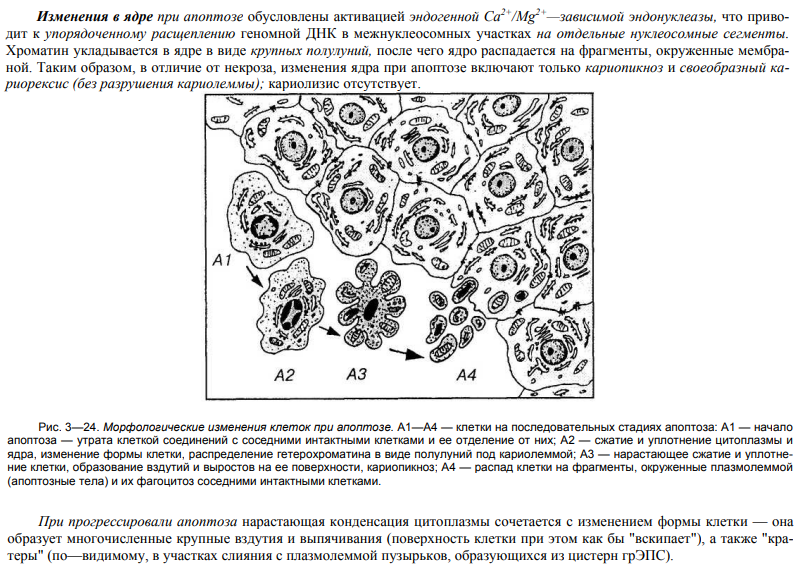
52. Структурно-функциональные изменения клеток при апоптозе.


![]()